


Abstract
Background Intervertebral disc degeneration is a major cause of low back pain. Previous researches have demonstrated local administration of signalling molecules as potential biological therapies for disc regeneration. Our laboratory has published encouraging results for effectiveness of injection of the cartilage derived morphogenetic protein-2 (CDMP-2) into ovine discs following annular injury. To elucidate the mechanisms underpinning these in vivo effects, this project aimed to investigate the potential of CDMP-2 on cellular migration, proliferation and extracellular matrix production in a human chondrocytic cell line.
Methods To evaluate cell motility, cells were seeded into Boyden chambers and CDMP-2 as a chemo-attractant or a stimulant was placed into either the bottom or top chambers respectively. Cells that had completed migration through the porous membrane were visualized by immunocytochemical staining and analysed using Image J. The effect of CDMP-2 on cell proliferation, proteoglycan and collagen production, as well as chondrogenic gene expression in human chondrocytic cell line C28/I2 was also examined.
Results The results revealed that cells migrated significantly under the influence of CDMP-2 (200 ng/ml) stimulation compared to control (3-fold increase, p = 0.033) and demonstrated a significant chemotactic movement towards a solution of 200ng/ml CDMP-2 (>2-fold increase, p = 0.027). A 35% increase in C28/I2 proliferation was observed after CDMP-2 stimulation (p < 0.0001) compared to control, and in the presence of 100ng/ml CDMP-2, proteoglycan synthesis had an 8-fold increase (p = 0.048). Similarly, gene expression analysis demonstrated increased expression of aggrecan, collagen types II, X and XXVII, BMPR-1A and BMPR-2 when cells were treated with CDMP-2.
Conclusion The study shows that C28/I2 cells can migrate under the influence of CDMP-2 as a chemoattractant or migration stimulator, suggestive of an effect on chondrocytic cells in the intervertebral disc. Further, CDMP-2 can stimulate C28/I2 cells to proliferate and synthesize key extracellular matrix proteins.
Background
Low back pain is one of the most reported chronic health problems in the adult population.1–3 It is rated the 6th most debilitating disease worldwide in terms of disability-adjusted life years (DALYs), and is the leading cause of disability in Australasia and Western Europe.4 Degenerative disc disease is one of the major causes of low back pain, with its prevalence estimated at 54% globally.5 The degenerative process of the intervertebral disc (IVD) is initiated by cell-mediated degradation of the nucleus pulposus (NP).6 The NP, which lies in the centre of the IVD with a meshwork matrix of proteoglycans, collagens and non-collagenous proteins, maintains disc hydration and mechanical integrity. These matrix components are produced in the NP by a heterogeneous group of notochordal cells, which dominate in early life, and chondrocyte-like cells, which dominate in adult life. It has been suggested that these chondrocyte-resembling cells are derived from the thin layer of cartilaginous endplate (EP) that is between the NP and the underlying bony vertebral body.7
Degenerative changes in the IVD increase the stress on surrounding tissues, and have been suggested to compress and/or sensitize nearby nerve fibres to manifest as low back pain.8, 9 The current standard for treatment of degenerative disc disease relies on pain relief, but does not address the mechanical or physiological repair of the disc.10 Hence, recent studies have explored biological strategies to reverse the pathological state of the cells and depleted extracellular matrix to a more physiological state. A particular strategy that has interested our group is the injection of signaling molecules into the disc to stimulate recruitment of healthy cells from surrounding tissues. Bone morphogenetic proteins (BMPs) are members of the TGF-β superfamily that have been of special interest in disc regeneration due to their endogenous potential for chondrogenesis.11
Cartilage-derived morphogenetic protein-2 (CDMP-2), otherwise known as growth differentiation factor-6 (GDF-6) or BMP-13, is an endogenous growth factor belonging to the BMP family and plays an important role in chondrocyte regulation.12, 13 In a comparative study of the effects of all known BMPs and CDMPs, increased collagen accumulation after adenoviral-mediated CDMP2 gene transfer was among the strongest of all BMPs tested, at 2.5-fold of control.14 However, the property that has distinguished CDMP-2 from other members of the same family (e.g. BMP-2) is its anti-osteogenic potential. A previous in vitro study highlighted that CDMP-2 inhibited osteogenic differentiation of human bone marrow mesenchymal stromal cells (BM-MSCs), while promoting chondrogenic differentiation, traits that may make CDMP-2 a more promising molecule for IVD therapy development than other better-known BMPs.15
In an in vivo study it was observed that injection of recombinant human (rh) CDMP-2 into ovine IVDs post annular injury resulted in enhanced cellularity in the disc NP tissues, with observable mobilization of cells from the cartilaginous EP to NP. This implied the possibility that CDMP-2 possessed chemotactic properties, attracting chondrocytic cells into the nucleus.16 To our knowledge, the migration of native disc cells induced by CDMP-2 has not been recorded previously, but a review of the literature suggests a strong possibility of CDMP-2 having such chemotactic properties.17 A demonstration of chemotactic ability in CDMP-2 implies that it would not only be therapeutic in mildly-degenerated discs, where the aim is to stimulate the metabolic activity of existing cells, but also in severe degeneration where the nucleus is depleted of cells. This would further enhance the potential of CDMP-2 to replenish the depleted NP and restore function to the degenerated IVD.
The aim of this project was to evaluate and characterise CDMP-2 induced cell migration in chondrocytic cells, as further evidence for the molecule's chemotactic potential. We hypothesised that the addition of CDMP-2 to chondrocytic cells, resembling those found in the cartilaginous EP of native discs, would increase cell migratory behaviour and elevate known chemotactic markers, as well as inducing proliferation and chondrogenic matrix production compared to unstimulated chondrocytes.
Materials And Methods
Cell culture
C28/I2 immortalized human chondrocytes, kindly donated by Dr Mary Goldring,18 were cultured in 75cm flasks in 1:1 Dulbecco's Modified Eagles Medium (DMEM) /F12 medium (Life technology, Carlsbad, CA, USA) with 10% (v/v) foetal calf serum (FCS; Life technology) and 1% (v/v) antibiotics-antimycotics (Life technology) at 37°C and 5% CO2 in an atmosphere of 95% air. The medium was changed every 48 hours. The cells were sub-cultured for experiments after growing to approximately 80–90% confluence.
Cellular proliferation assay
The effect of CDMP-2 on the proliferation of chondrocytes was assessed using the CelTitre96® Aqueous one solution proliferation assay (MTS assay, Promega, Madison, WI, USA) as per manufacturer's instructions.
To determine the optimal dose response of CDMP-2 on cell proliferation, 100µl of cells (0.5 x 105 cells/ ml) were seeded into 96-well plates and cultured for 24 hours. The medium were replaced with 200µl of growth medium containing CDMP-2 (Recombinant human CDMP-2, PeproTech, Rocky Hill, NJ, USA) at 0ng/ml, 50ng/ml, 100ng/ml, 150ng/ml, or 200ng/ ml. The plates were incubated for 48 hours, and then 20µl of MTS reagent was added to each well. The plates were incubated for a further 2 hours, and measured for absorbance at a wavelength of 490 nm in a spectrophotometer.
To determine the optimal time points of CDMP-2 on cellular proliferation, cells were seeded as described above, and then treated with 100µl of either the growth medium alone or with 100 ng/ml CDMP-2. The plates were incubated for 24, 48 or 72 hours, at the end of which MTS reagent was added and the absorbance was measured at 490nm in the spectrophotometer. Each experiment was conducted with five different cell culture samples (n = 5).
Cell migration
Migration of C28/I2 cells in vitro was detected using the Boyden chamber assay. To achieve this, cultured cells were treated with the migration medium, consisting of DMEM/F12 medium with 1%FCS. Cells in 100µl of migration medium at 1x105 cells/ml were seeded in the top chamber inserts (8nm pores) of a 24-well Boyden-Chamber plate (Becton and Dickinson, Franklin Lakes, NJ) and cultured in the incubator for 1 hour before being divided for two experiments: chemo-stimulation and chemo-attraction. For chemo-stimulation, cells were incubated for 1hr, then 100µl of migration medium containing 0ng/ml, 100ng/ml, 200ng/ml or 300ng/ml of CDMP-2 was added into the inserts with the seeded cells; 600µl of migration medium was then added to the bottom wells and the plates were cultured for 48 hours. For chemo-attraction, 100µl of additional migration medium was added to the seeded cells in the top chambers. In the bottom wells, 600µl of migration medium containing 0ng/ml, 100ng/ml, 200ng/ml or 300ng/ml of CDMP-2 was added. Plates were then cultured at 37°C under 5%CO2 for 48 hours. After the 48-hour period, the inserts were washed with PBS and cells were fixed with 4% paraformaldehyde for 10 minutes, at the end of which the inserts were washed with PBS. The inserts were inverted and the membranes were stained with 0.2% Crystal Violet solution (Sigma), visualising the cells on the underside of the membrane (migrated cells). The cells adhering to the top of the membrane (non-migrating cells) were scraped off using cotton swabs. The membranes were air dried, mounted onto glass microscope slides and enclosed with coverslips. Slides were viewed under a Leica light microscope with a digital camera, and 3 images of random locations in each slide were photographed at 10x magnification. Each experiment was duplicated, with total images analyzed being n = 6.
Alcian blue staining
Proteoglycan (PG) production was estimated by Alcian blue staining. Briefly, 1ml of cells (0.5 x 105 cells/ml) were seeded into 6-well plates with glass cover slips in each well and incubated for 24 hours. Cells were then treated with or without 100 ng/ml CDMP-2 in growth medium for 48 hours. After wash with PBS, cells were fixed with 4% paraformaldehyde prior to staining with 1% Alcian blue solution (Sigma-Aldrich, St. Louis, MO, USA) in 3% acetic acid for 30min. The cells were washed with water, air-dried and then examined using a Leica light microscope. Images of 5 random fields in each cover slip were photographed with a Leica camera. Captured images were semi-quantitatively analysed using the software ImageJ to determine the amount of PG present, which was highlighted by the Alcian blue stain. Each experiment was conducted in triplicate (n = 3).
Image analysis
Photographs were saved as jpg files and then opened with ImageJ software (version 1.47i, National Institute of Health, Bethesda, MD, http://rsbweb.nih.gov/ij/). Images were analysed with methodology as highlighted by Gutierrez and colleagues [19].
[3H]-proline incorporation assay
Total collagen synthesis was assayed by measurement of cellular [3H]-proline incorporation. Briefly, cultures were treated with or without CDMP-2 (100ng/ml) for 48h and then 10µCi/ml [3H]-proline (Perkin-Elmer, Life Sciences, Boston, MA) was added to each well for further 12 hours incubation. At the end, cells were washed 3 times with cold PBS and incubated with 5% trichloroacetic acid (TCA) for 30 min. After 2 rinses with cold 5% TCA, the acid-precipitate material was solubilized overnight with 0.5ml of 0.5 M NaOH at 37°C. Finally, all reactions were stopped by adding 0.5 M HCl, and harvested materials were placed in 2ml liquid scintillation and measured in a β-counter (TL 5,000 sec, Beckman Instruments, Inc., Fullerton, CA). The resulting value of collagen synthesis were normalized by total protein and expressed as cpm/ug of protein.
Immunocytochemical analysis
Immunocytochemistry was used to observe the functional expression of chondrocytic proteins in C28/I2 cells following CDMP-2 stimulation. 1ml of cells (0.5 x 105 cells/ml) was seeded into 6-well plates with glass cover slips in each well and incubated for 1 hour. Cells were then stimulated with CDMP-2 (0ng/ml, 100ng/ml, 200ng/ml) in growth medium for 48 hours. At the end of culture, cells were fixed with 5% paraformaldehyde for 15 min and washed with PBS. Endogenous peroxide activity was depleted with 3% hydrogen peroxide (Sigma-Aldrich, St. Louis, MO) for 30 min before blocking for nonspecific binding with 1% bovine serum albumin (BSA) for 15 min. Cells were then incubated with goat anti-collagen-2 polyclonal (1:100; Santa Cruz Biotechnology, Santa Cruz, CA), or mouse anti-aggrecan monoclonal (1:150; Chemicon International, Temecula, CA) primary antibodies for 2 hours. The wells were washed with 0.1% Triton-X100 in PBS and incubated with MULTILINK solution (DAKO, Botany, Australia) for 15 min before application of streptavidin-conjugated peroxidase incubation for 20 min. The results were visualized with 3,3’-Diaminobenzidine solution (DAKO). Negative controls received identical treatments apart from the omission of the primary antibody. Wells were imaged under a Leica light microscope with a digital camera (Leica DC200, GmbH, Wetzlar, Germany). Five images were captured at random locations in each well and images were analysed using ImageJ in a similar manner to the Alcian blue assay.
Real-time RT-PCR
C28/I2 cells were cultured in 25cm flasks for 48 hours with CDMP-2 stimulated growth media (concentrations of 0ng/ml, 100ng/ml and 200ng/ml).
Following culture, total RNA was isolated from the cells using H-RNA purification kits (Roche Diagnostics, Manheim, Germany) as described by the manufacturer. cDNA was generated by reverse transcription of 1µg of total RNA using SuperScript III first-strand synthesis kit (Invitrogen). The cDNA was used in 20µl reactions for real-time PCR analysis using a Rotor-Gene RG3000 system (Corbett Life Science, Sydney). The thermal profile for all reactions was as follows: 5 min at 95°C, followed by 40 amplification cycles of 15 sec at 95°C, 30 sec at 60°C and 15 sec at 72°C. All primers were designed from published mRNA sequences. The cycle threshold (Ct) value for each reaction as determined, and the relative mRNA expression of various genes was calculated using the Comparative CT Method with the average Ct value of three housekeeping genes- glycer-aldehyde-3-phosphate (GAPDH), beta-2-microglobulin (B2M) and hypoxanthine phosphoribosyl-transferase I (HPRT1). The reaction for each gene was repeated four times.
Statistical method
Statistical analysis was performed using SPPI software (Systat Software Inc., Chicago, IL). The data is expressed as mean ± standard deviation (SD). Normally distributed data was analysed by Student's T-Test and One-way Analysis of Variance (ANOVA). A p-value < 0.05 was considered significant.
Results
CDMP-2 stimulates cell proliferation
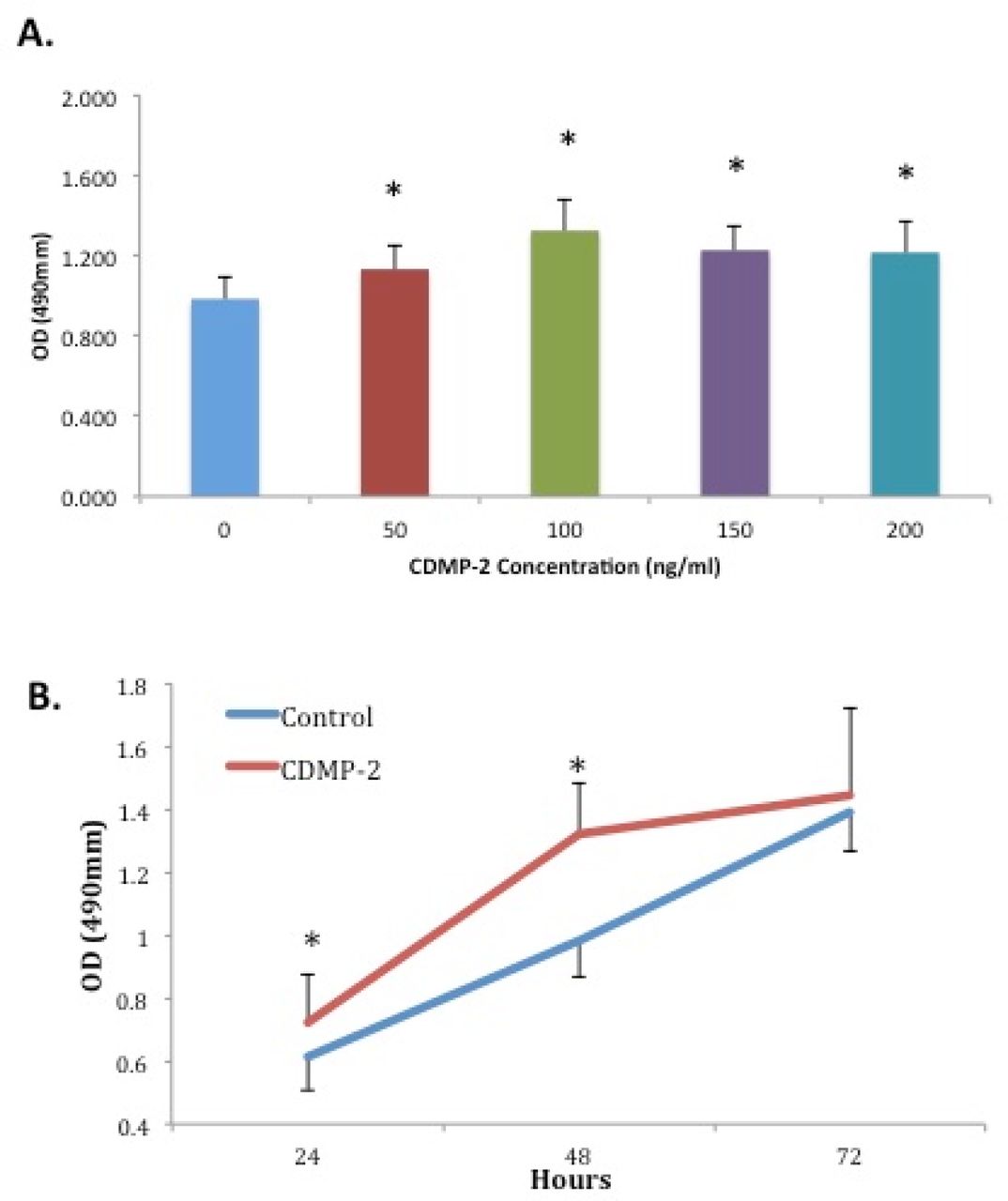
C28/I2 cells incubated for 48 hours with culture medium and varying doses of CDMP-2 were analysed using MTS assays. All groups treated with CDMP-2 demonstrated significantly increased absorbance (p < 0.05) compared to control (Figure 1A). In particular, cells treated with 100ng/ml of CDMP-2 yielded the highest absorbance, with a 35% increase compared to control (p < 0.0001). Cells treated with 50ng/ml of CDMP-2 yielded the lowest absorbance with a 16% increase (p = 0.001). The concentration of 100ng/ml of CDMP-2 was determined as optimal for the stimulation of cell proliferation and was used for further proliferation and cellular production experiments.

A) Effect of CDMP-2 on cellular proliferation of C28/I2 cells at varying doses for 48 hours. Absorbance was significantly higher (p < 0.05) in all groups treated with CDMP-2 compared to control. Each bar represents combined data from three individual experiments performed in quintuplicate (mean±SD). * indicates significant difference compared with control group (p < 0.05). B) Effect of CDMP-2 (100ng/ml) on cellular proliferation of C28/I2 cells at varying times. At 24 and 48 hours, there was significantly higher absorbance (p< 0.05) in groups treated with CDMP-2. Data is presented as mean ± SD and results are derived from three individual experiments each performed in quintuplicate. * indicates significant difference compared with the control group of the same time point (p < 0.05).
In determining the proliferation response over time, both the control and CDMP-2 (100ng/ml) groups of cells were cultured for 3 time-points and compared for absorbance. In both the control and CDMP-2 groups, an increase in time was associated with an increase in absorbance (Figure 1B). At 24 hours, CDMP-2 treated cells showed an 18% increase in absorbance compared to control (p = 0.046). Increased absorbance was most prominent at 48 hours, with the CDMP-2 group having 35% more absorbance than control (p < 0.0001). There was no significant difference in absorbance at 72 hours when compared to the control (p = 0.476) (Figure 1B).
CDMP-2 stimulates chondrocyte migration
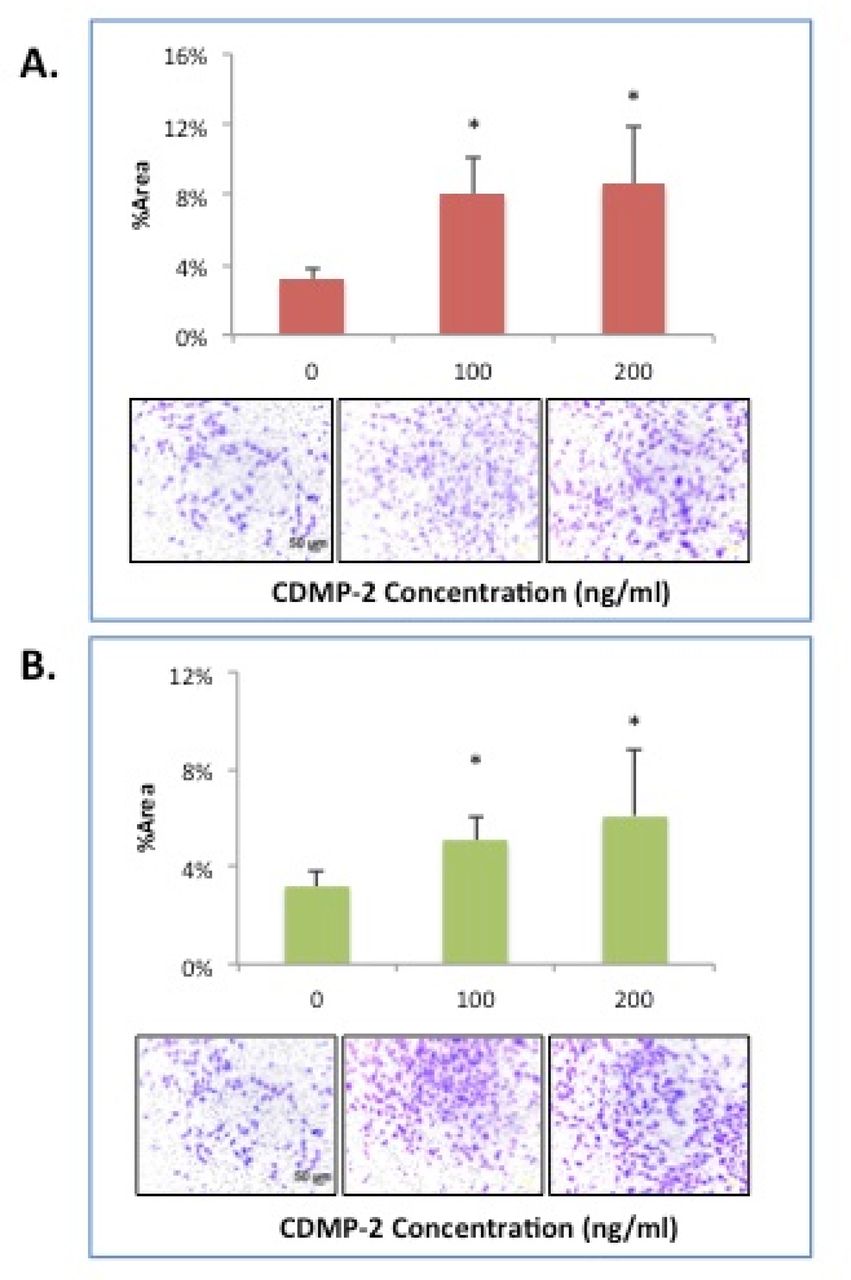
For migration experiments, 100ng/ml and 200ng/ml of CDMP-2 were used in comparison to control to establish any dose-response patterns. When CDMP-2 was used as a stimulant, there were significant increases in the mean %area stained on the underside of Boyden chamber membranes at all concentrations (n = 6, p < 0.05), indicative of increased cell migration. Cells stimulated with 200ng/ml of CDMP-2 yielded the highest, a ~3-fold increase (p = 0.013) in %area compared to the control (Figure 2A&B). There was a positive trend between %area and CDMP-2 concentration as a stimulant, indicative of a dose response.

A) Effect of CDMP-2 on C28/CI2 cell migration capacity. The presence of CDMP-2 in the upper chamber of a transwell plate significantly increased cell migration as observed with Boyden Chamber assay with varying doses at 45 hours, compared to cultured alone. (Top panel) Graphical representation of Crystal Violet –stained cells after migration; percentage of area staining was calculated with Image-J software. (Bottom panel) Representative microscopic images of Boyden Chamber filters mounted on slides. Varying concentrations of CDMP-2 were used. All groups with CDMP-2 as a stimulant had significantly higher %area of staining compared to control (p < 0.05). Data is presented as mean ± standard deviation (SD). * indicates significant difference compared with control (p < 0.05). B) Effect of CDMP-2 on C28/CI2 cell migration capacity. The presence of CDMP-2 in the bottom chamber of a transwell plate significantly increased cell migration as observed with Boyden Chamber assay with varying doses at 45 hours, compared to cultured alone. (Tope panel) Graphical representation of Crystal Violet –stained cells after migration; percentage of area staining was calculated with Image-J software. (Bottom panel) Representative microscopic images of Boyden Chamber filters mounted on slides. Varying concentrations of CDMP-2 were used. As an attractant, doses of 200ng/ml and 300ng/ml of CDMP-2 resulted in significantly higher % area of staining compared to control (p < 0.05). Data is presented as mean ± standard deviation (SD). * indicates significant difference compared with control (p < 0.05).
When CDMP-2 was used as an attractant, the highest %area was observed at 200ng/ml with a >2-fold increase compared to the control (p = 0.027) (Figure 2B), with the results again demonstrating a dose-response trend. However, CDMP-2 as an attractant yielded less migration than when present as a stimulant at all concentrations (Figure 2A& B).
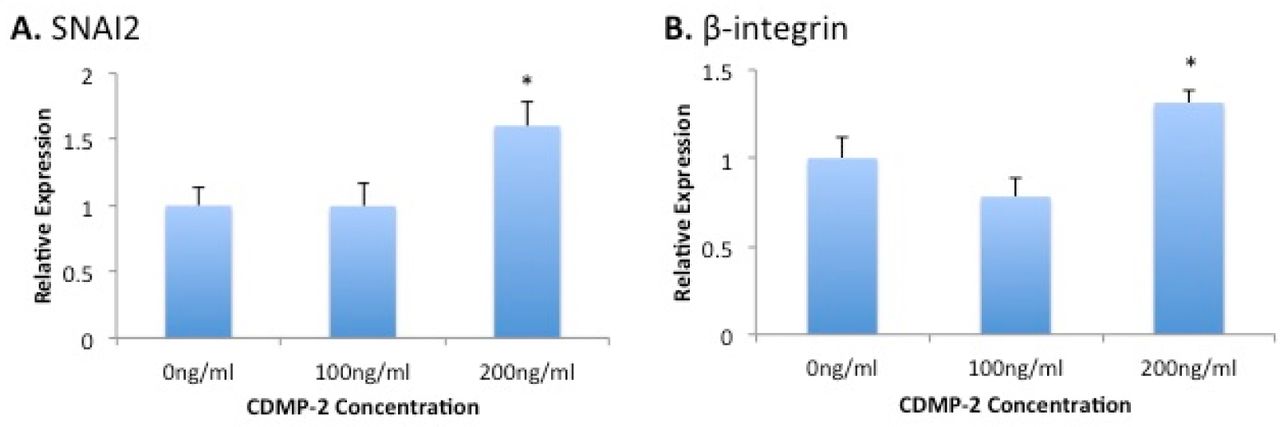
To investigate the potential involvement of known migration marker genes, we analysed the effect of mRNA expression of the cells following culture with CDMP-2. We observed significant increases in expression of SNAI2 (60% increase, p = 0.051) and β-integrin (30% increase, p = 0.047) mRNA when cells were cultured with 200ng/ml of CDMP-2, compared to the control. However, no significant differences in the expression of these markers were observed at 100ng/ml of CDMP-2 (Figure 3).
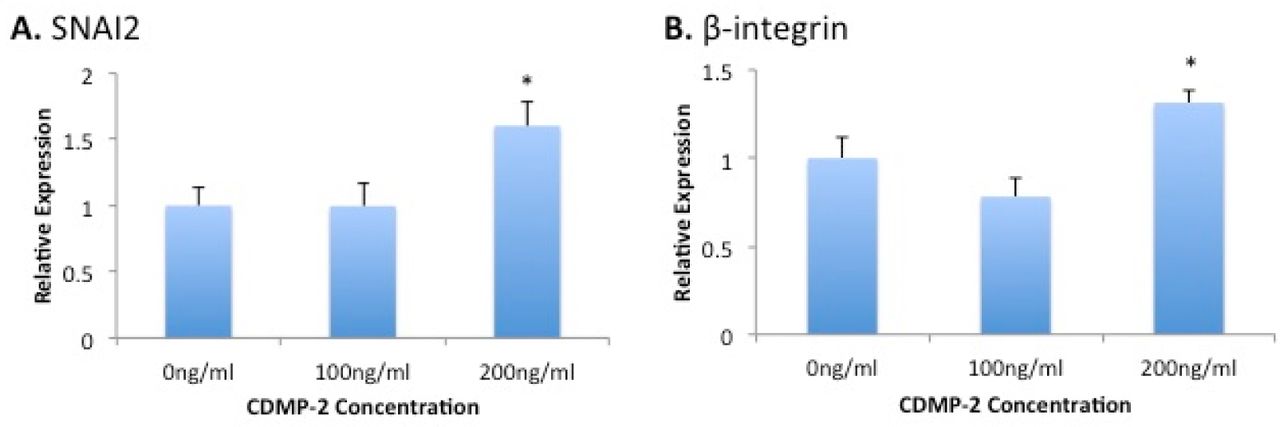
Gene expression analysis of migration markers in C28/I2 cells under CDMP-2 stimulation. Represented as CDMP-2 concentration (x-axis) against normalised relative expression (y-axis). Relative expression was calculated using RT-PCR analysis, with 3 reference genes (GAPDH, B2M, HPRT1). Values were normalised to the control (0ng/ml). Each reaction was repeated 4-times, values represented as mean + SEM. * indicates significant difference compared to control group (p < 0.05). A) SNAI2; B) β-integrin.
CDMP-2 stimulates proteoglycan production
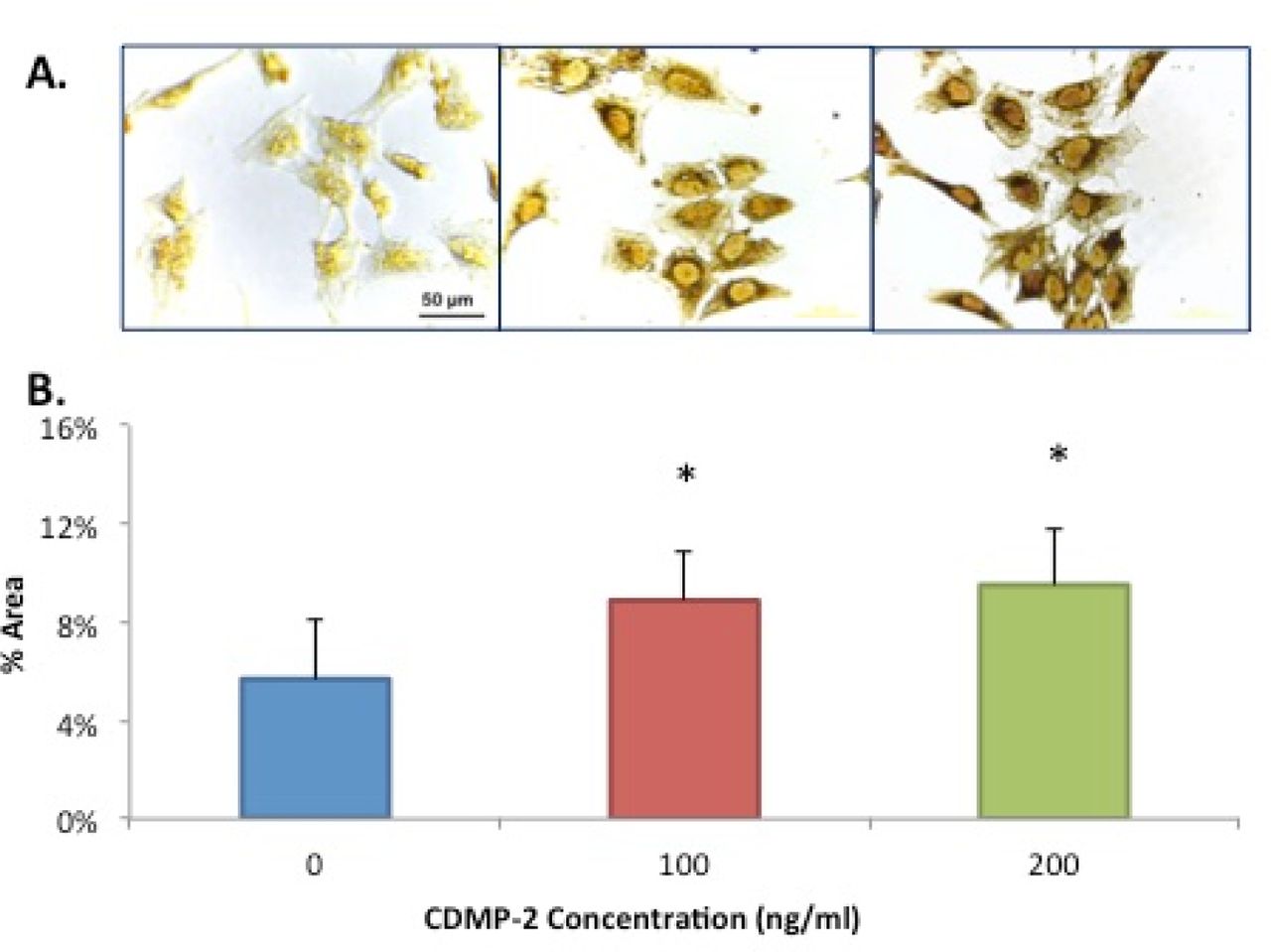
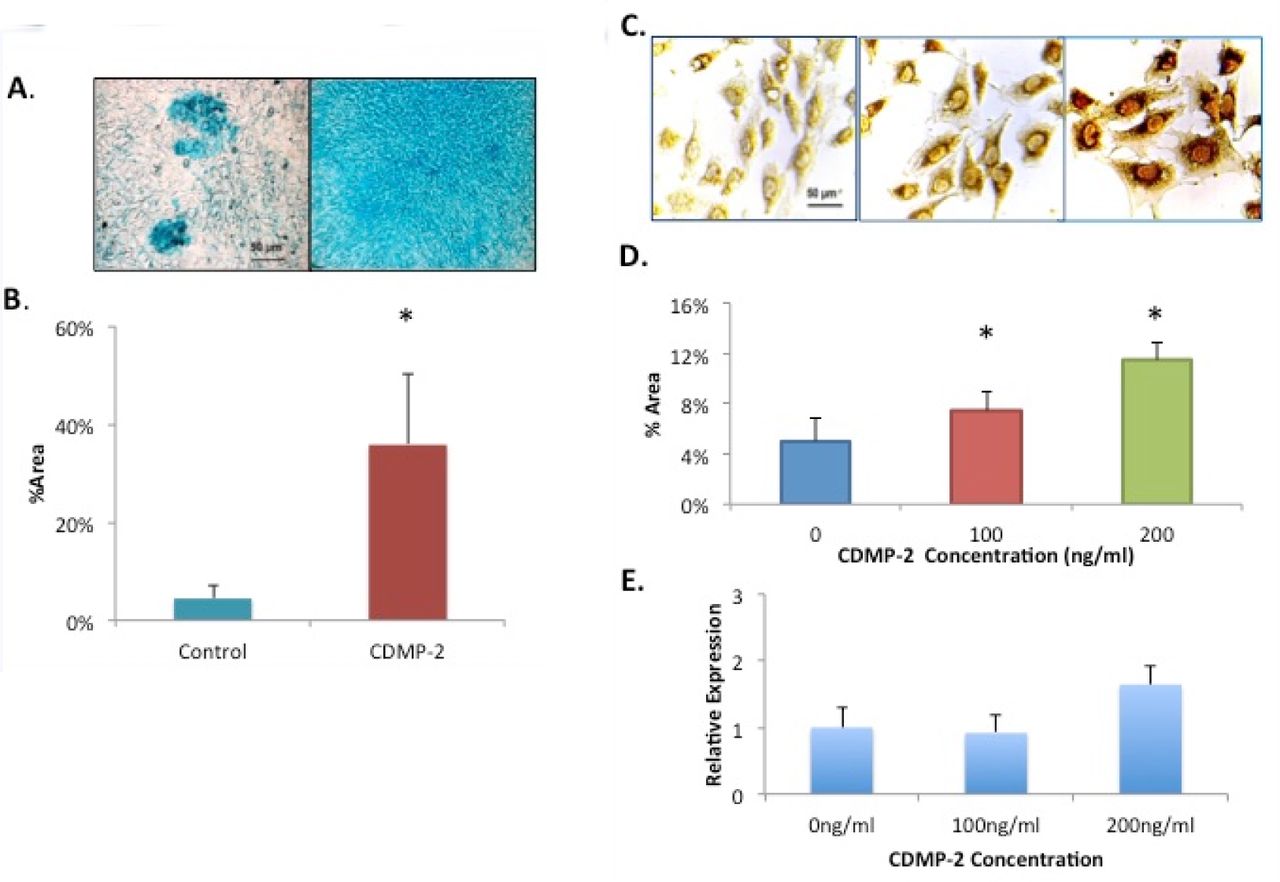
Alcian blue staining and ImageJ analysis were used to determine total PGs production in C28/I2 cells cultured with CDMP-2 (100ng/ml). A significantly (n = 5, p = 0.048) higher percentage area of positive staining was detected (Figure 4A), with an almost 8-fold increase compared to the control (Figure 4B).

A) Microscopic view of C28/I2 cells with Alcian blue staining. Stained cells with GAG production is observed to be more prominent in the group treated with CDMP-2 (right) than the control group (left). B) Effect of CDMP-2 (100ng/ml) on proteoglycan production as observed with Alcian blue staining assay and ImageJ analysis. Mean %area of staining is 7.8 times more in the CDMP-2 treated group than the control group (p = 0.048). Data is presented as mean ± standard deviation (SD) with each bar representing the combined data from three individual experiments. C) Microscopic view of C28/I2 cells of immuno-staining with primary antibody for aggrecan. Strong brown staining is observed in cells stimulated with CDMP-2 of 100ng/ml (middle panel) and 200ng/ml (right panel) than with control (left panel). The figure is a representative of cultured from three independent samples (original magnification: X400). D) Mean % area of staining is used to quantify aggrecan immuno-staining of C28/I2 cells. Under the influence of CDMP-2 of varying concentrations, cells increased functional production of aggrecans by 50% at 100ng/ml and 130% at 200ng/ml compared to the control (p < 0.05). Data is presented as mean ± standard deviation (SD). E) Gene expression analysis of aggrecan in C28/ I2 cells under CDMP-2 stimulation. Represented as CDMP-2 concentration (x-axis) against normalised relative expression (y-axis). Relative expression was calculated using RT-PCR analysis, with 3 reference genes (GAPDH, B2M and HPRT1). Values were normalised to the control (0ng/ml). Each reaction was repeated 4-times, values represented as mean + SEM. Results were not statistically significant (p < 0.05). * indicates significant difference compared with the control group (p < 0.05).
Immunocytochemical staining of C28/I2 cells was used to model CDMP-2 stimulation of functional aggrecan protein expression. Compared to the control (0ng/ml), there was a 50% increase in the %area of staining at 100ng/ml CDMP-2 (n = 5, p = 0.021) and a 130% increase with a 200ng/ml dose (n = 5, p = 0.003) (Figure 4C&D). Two doses were used in this experiment to establish a dose-response. We also analysed the expression of aggrecan mRNA in CDMP-2 stimulated C28/I2 cells using semi-quantitative PCR, detecting an increase of 60% in aggrecan expression relative to housekeeping genes at 200ng/ml of CDMP-2 when compared to the control (0ng/ml CDMP-2) (data not shown). The increased expression of aggrecan demonstrated a dose-related pattern (Figure 4E).
CDMP-2 stimulates collagen expression and production
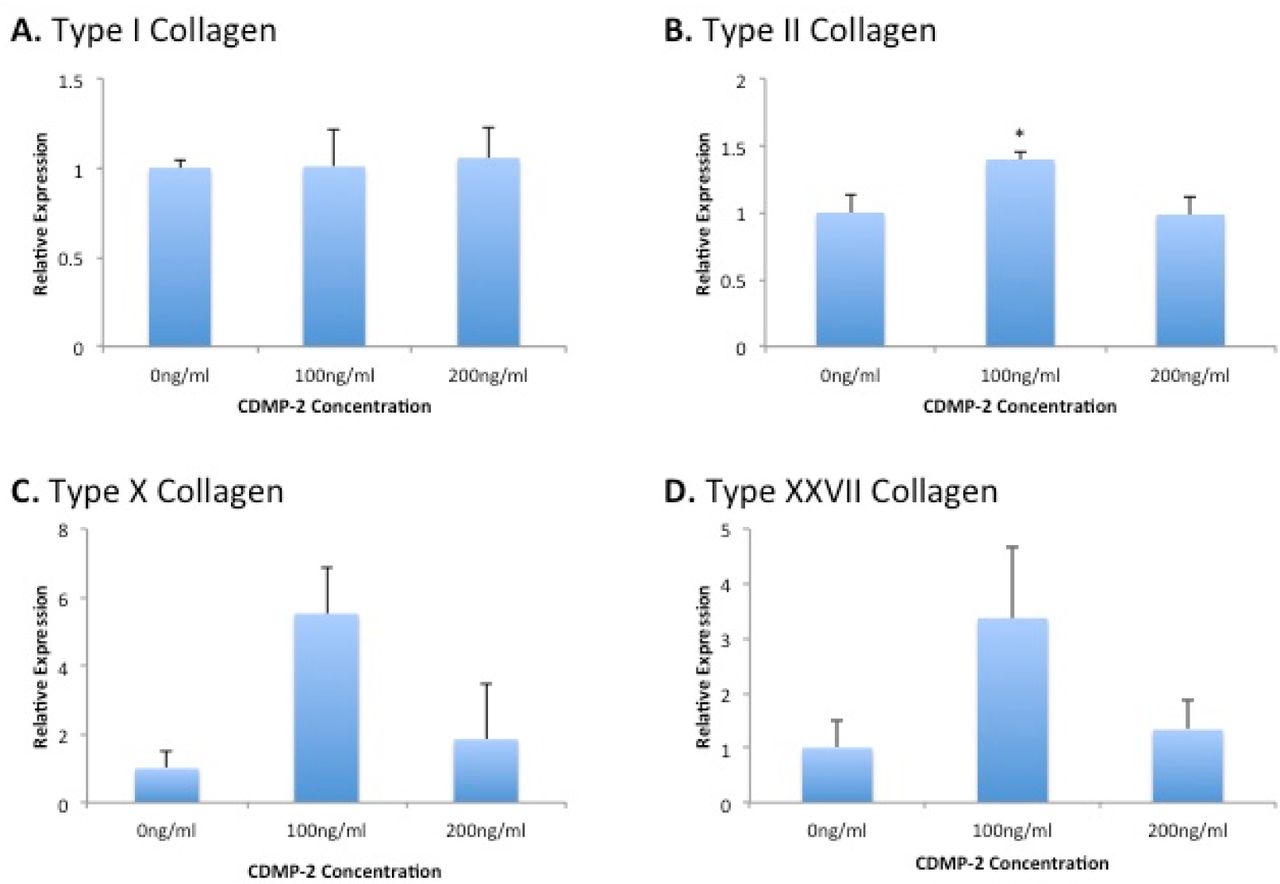
As a further indicator of chondrogenic potential, we analysed the production of collagen by C28/I2 cells in response to CDMP-2. Total collagen synthesis was measured using [3H]-proline incorporation assay of cells cultured with/without 100ng/ml of CDMP-2 for 48 hours. There was a 3-fold increase (p = 0.014) in collagen synthesis when cells were treated with CDMP-2 compared to cells treated with growth medium alone (Figure 5). This is supported by gene expression analysis of various collagens in CDMP-2 treated C28/I2 cells where stimulation with 100ng/ ml of CDMP-2 caused a 40% increase in expression of type II collagen (p = 0.018), compared to the control (Figure 6). There was also a 5.5-fold increase of type X collagen (p = 0.099) and 3.5-fold increase of type XXVII collagen (p = 0.254) at this concentration of CDMP-2, though the results were not statistically significant.

Effect of CDMP-2 (100ng/ml) on total collagen production measured with [3H]-proline incorporation assay. Total collagen synthesis is 3 times more in the CDMP-2 treated group than the control group (p < 0.05). Data is presented as mean ± standard deviation (SD) with each bar representing combined data from three individual experiments.

Gene expression analysis of chondrocytic markers and receptors in C28/I2 cells under CDMP-2 stimulation (0ng/ml, 100ng/ml, 200ng/ml) for 48 hours. Represented as CDMP-2 concentration (x-axis) against normalised relative expression (y-axis). Relative expression was calculated using RT-PCR analysis, with 3 reference genes. Values were normalised to the control (0ng/ml). Each reaction was repeated 4-times, values represented as mean + SEM. * indicates significant difference compared with the control group (p < 0.05). A) Type I Collagen; B) Type II Collagen; C) Type X Collagen; D) Type XXVII Collagen.
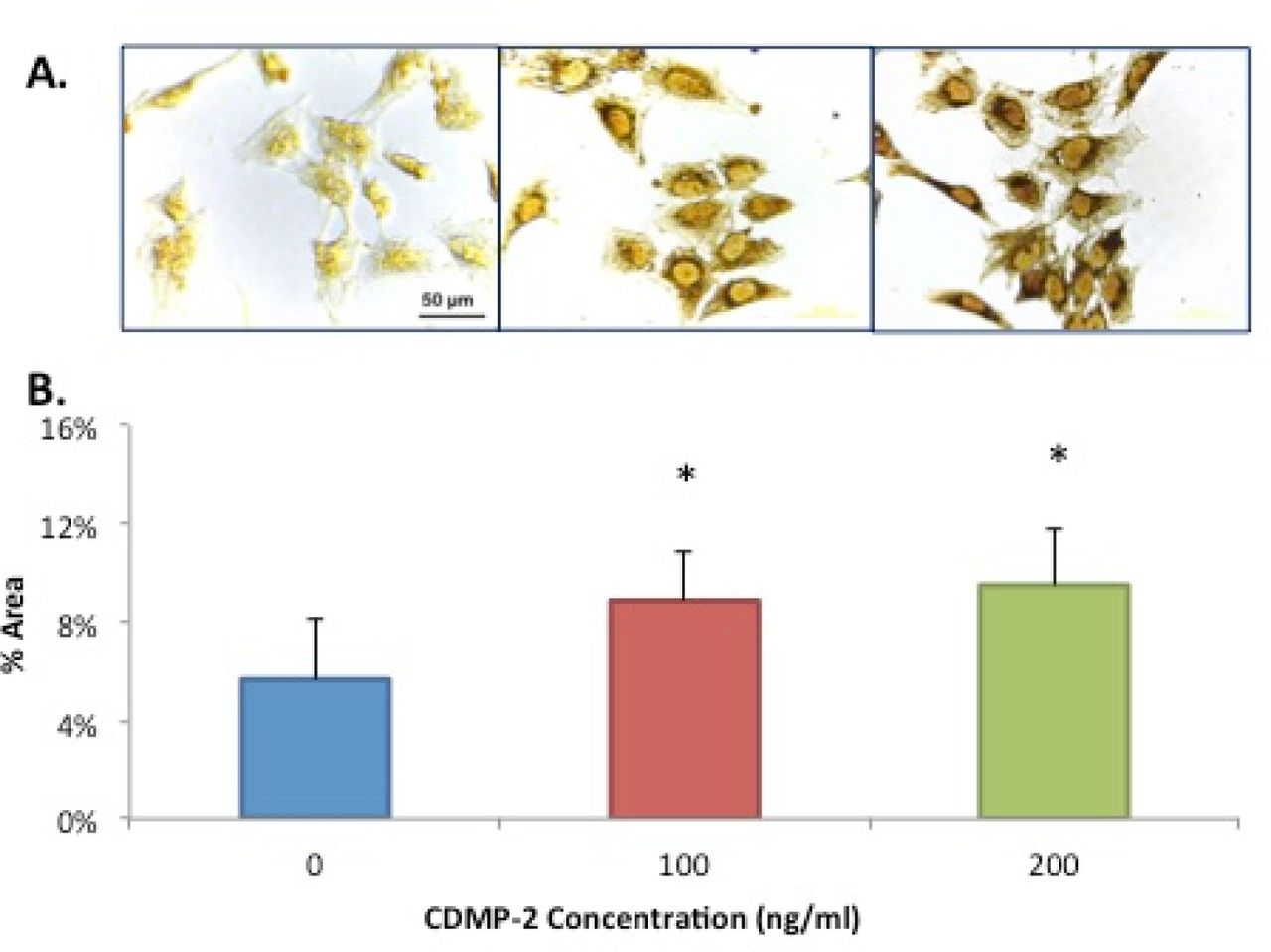
Likewise, immunocytochemistry of C28/I2 cells with primary antibody for type II collagen suggested a dose dependent increase in expression of type II collagen protein with CDMP-2 stimulation. A 100ng/ml dose of CDMP-2 induced a 60% increase in %area of staining (n = 5, p = 0.023) compared to control (Figure 7A). We further investigated with a 200ng/ml dose, which caused a 70% increase (n = 5, p = 0.002) (Figure 7B). No brown staining was detected in the negative control slides with the omission of the primary antibody (results not shown). However, when we performed PCR to confirm this finding, it was observed that at 200ng/ml of CDMP-2, type II collagen expression was not significantly different from that of control (Figure 6B)
A) Microscopic view (x40 magnification) of C28/I2 cells of immuno-staining for type II collagen. Strong brown staining is observed in cells stimulated with CDMP-2 of 200ng/ml (right panel) and 100ng/ml (middle panel) than with control (left panel). The figure is a representative of culture from three independent samples. B) Protein expression of type II collagen production of C28/I2 cells under CDMP-2 influence, demonstrated using immunocytochemistry and ImageJ analysis. Data presented as mean ± standard deviation (SD). N = 5. * indicates significant difference compared with control (p < 0.05).
BMP receptor expression in CDMP-2 treated chondrocytic cells
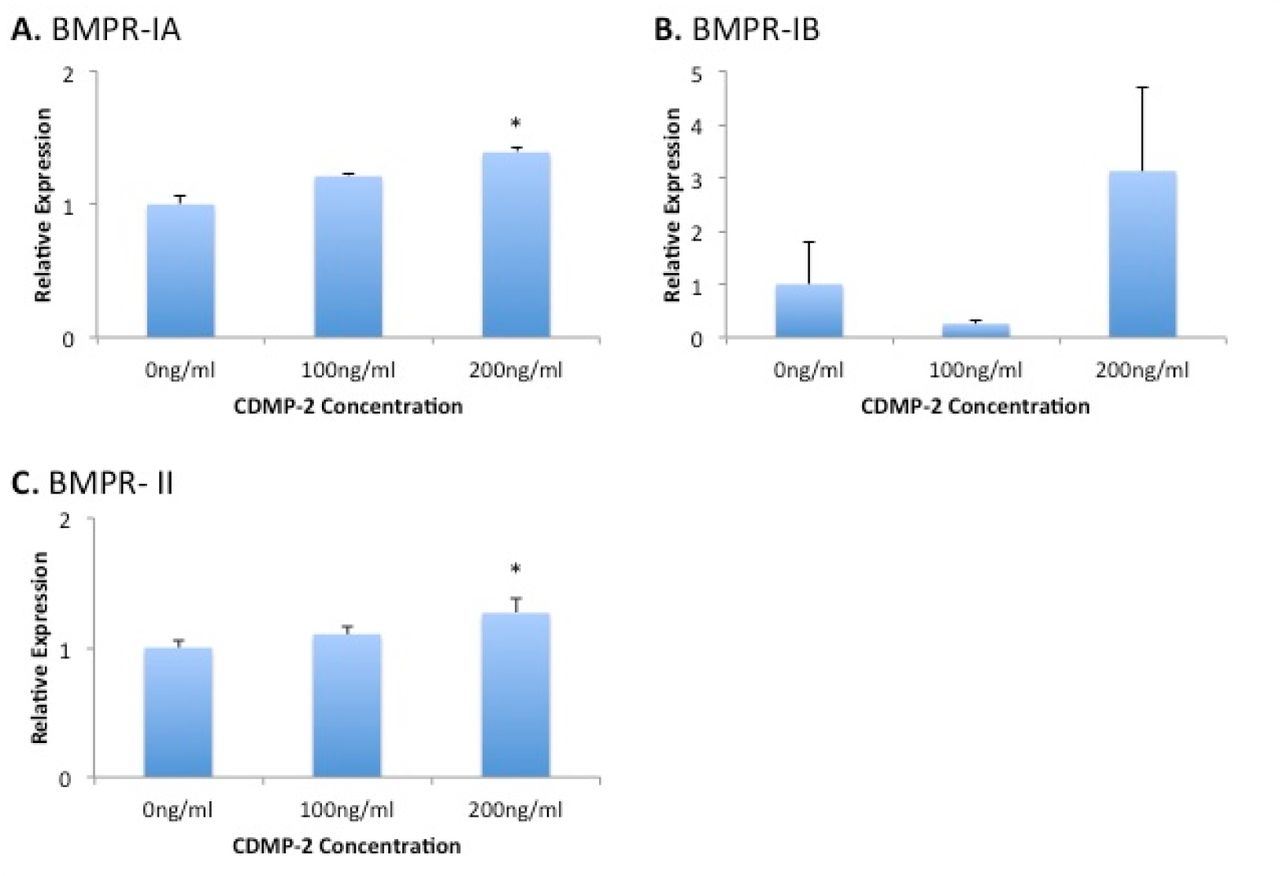
To further investigate the mechanism of CDMP-2 chondrogenic action, we measured the gene expression of related receptors in CDMP-2 stimulated C28/I2 cells at doses of 100ng/ml and 200ng/ml. Stimulation with CDMP-2 at 200ng/ml had statistically significant effects on the expression of BM-PR-1A and BMPR-II, with a 40% (p = 0.02) and 30% (p = 0.02) increase from the control respectively (Figure 8). BMPR-IB expression was increased in response to CDMP-2 stimulation at 200ng/ml, but the data was more variable (Figure 5).
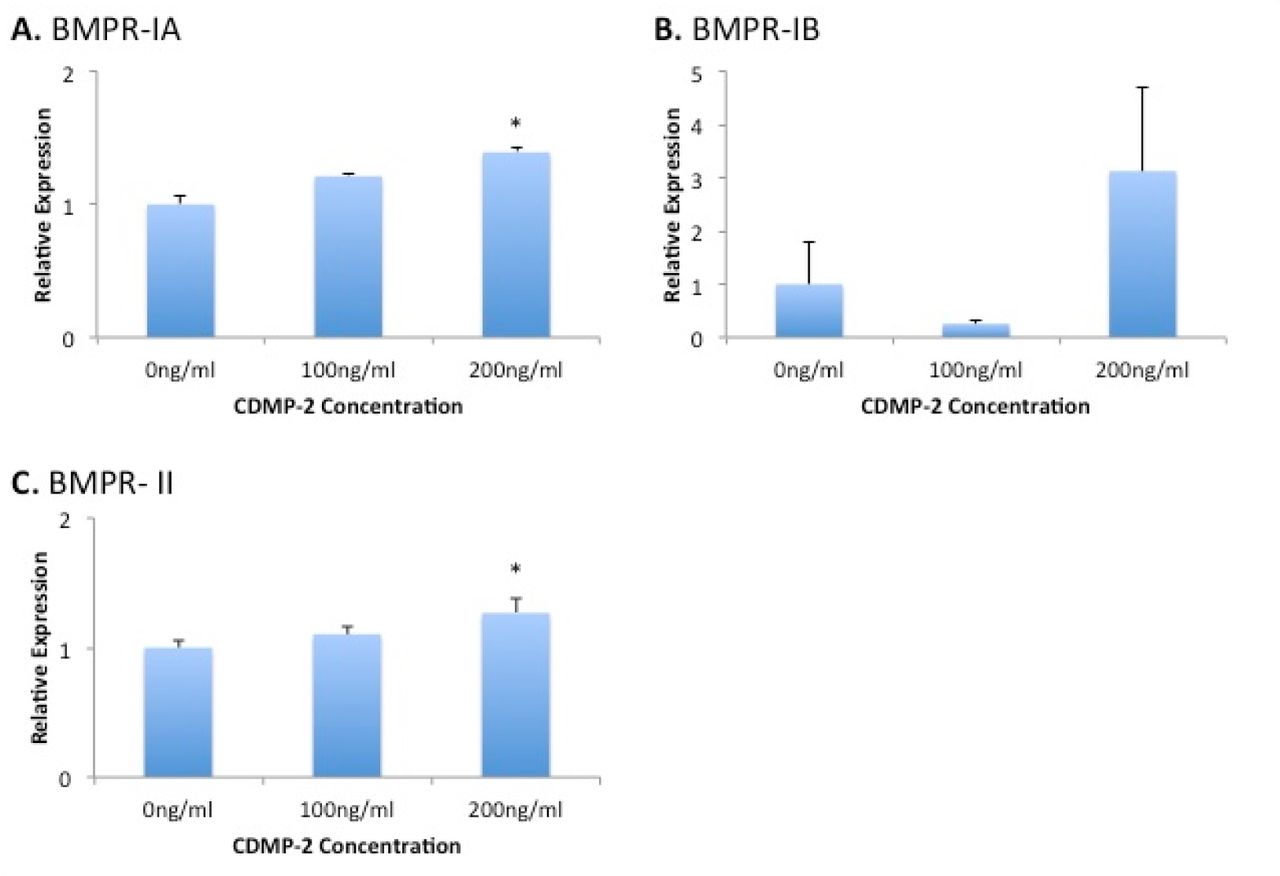
Gene expression analysis of BMP receptors in C28/I2 cells under CDMP-2 stimulation. Represented as CDMP-2 concentration (x-axis) against normalised relative expression (y-axis). Relative expression was calculated using RT-PCR analysis, with 3 reference genes. Values were normalised to the control (0ng/ml). Each reaction was repeated 4-times, values represented as mean + SEM. * indicates significant difference compared to control group (p < 0.05). A) BMPR-IA; B) BMPR-IB; C) BMPR- II.
Discussion
This study demonstrates for the first time the ability of CDMP-2 to induce cell migration in a human chondrocyte cell line. The data support previous observations of enhanced cellularity at the EP-NP junction following intra-discal CDMP-2 injection in an ovine injury model and the hypothesis that cells were moving towards the NP.16 The decision of employing an immortalised cell line rather than native IVD cells for this study was based on the faster cellular turnover and reduced genetic variability of the C28/ I2 cells, which have been previously used to study cartilage metabolism.18 This suits our study objective to establish the biological ability of CDMP-2 to induce cell movement, in cells which resemble those of the cartilaginous endplate, with the benefit of reduced individual variability inherent in any primary cell based study. Clearly the conclusions cannot be directly extrapolated to apply to primary cells, however with the principle established, further studies on the CDMP-2 induced motility of disc cells and mesenchymal progenitors is planned and will answer questions as to cell type specificity in CDMP-2 migratory response behaviour.
The current study shows that CDMP-2 indeed has the ability to induce cell motility, at least in the chondrocytic C28/I2 cell line. The Boyden chamber assays quantified cell movement in response to CDMP-2 when cells were stimulated with CDMP-2 (stimulant, 2-3 fold) or induced to move along a CDMP-2 gradient (attractant, 2-fold). These data are in keeping with previous reports of chemotactic effects in BMPs such as BMP-2, a known chemotaxis-inducing growth factor.20
Interestingly, we observed a greater and more dose-dependent migration response when CDMP-2 was employed as a chemo-stimulant than a chemo-attractant. Two key processes, cell polarisation and gradient sensing, influence the pseudopodia formation and motility of the cells.21, 22 The results here suggest that the migration of the current cell line may be more dependent on close-range CDMP-2 stimulation driving the cells to polarise than a long-distance attractant gradient, but further studies are needed to establish the mechanism of action. Such properties become significant when considering any future therapeutic use of CDMP-2 in the disc, as the injection of the molecule may be more effective in the specific location of the native cells (e.g. cartilaginous EP) than the desired final destination of the migrating cells (NP). It is also possible that responses to CDMP-2 may vary depending on the cell type. It will be particularly interesting to establish the migration of mesenchymal progenitors such as those identified in disc stem cell ‘niches,’23, 24 in response to CDMP-2. The observed cell migration in response to CDMP-2 was accompanied by up-regulated expression of SNAI2 and β-integrin genes. Expression of human SLUG (SNAI2) gene correlates with increased invasiveness and metastases of tumours,25 while adhesion molecule beta-integrin is known to facilitate intracellular signalling in the migration of cancer cells.26 In this study, expression of these migration markers was determined in static cell cultures stimulated with CDMP-2, rather than in migrated cells. It would be interesting in future studies to develop a migratory cell model with which to study gene expression in CDMP-2-induced cell migration.
CDMP-2 has been suggested to regulate chondrocyte differentiation and proliferation,27 particularly in tissues delineating bone-cartilage borders.28, 29 In this study we examined the effects of CDMP-2 incorporation on cellular proliferation and extracellular matrix production in a human chondrocytic cell line in vitro. The MTS assay results showed that CDMP-2 did induce cellular proliferation of C28/I2 cells (increase by 35%) and, though limited dose response trends were observed, the proliferative effect was present through to 72 hours of stimulation. Recent data demonstrating the proliferative potential of CDMP-2 on murine MSCs15, 30 and CDMP-2 expression in proliferating cartilaginous tissue31 support these data, with mouse MSCs proliferating in response to CDMP-2 for as long as 168 hours.15
CDMP-2 was observed to have a functional chondrogenic effect, showing stimulation of proteoglycan and collagen synthesis in C28/I2 cells. The total glycosaminoglycan (GAG) content reflected in Alcian blue staining showed that cells readily produced more proteoglycans under the influence of CDMP-2, and more specifically CDMP-2 stimulation elevated aggrecan protein and mRNA levels, an important proteoglycan in disc maintenance. These data are reinforced by other investigations in our laboratory, which have demonstrated similar patterns of GAG production in human IVD cells when cultured with CDMP-2.32 This, conversely, validates our study, as the results derived from the cell line appear to resemble the effect CDMP-2 has on endogenous disc cells. Our results are further supported by previous reports of enhanced proteoglycan production in response to in vivo CDMP-2 injection into injured ovine IVD. 16 CDMP-2 has also reportedly enhanced mRNA levels of proteoglycans and aggrecans in mouse chondrocytic cell line MC615.33 As degenerative disc is characterized by a loss of PG, it is important and encouraging that CDMP-2 has such a potent effect on expression of these IVD extracellular matrix molecules.
CDMP-2 stimulation increased synthesis of total collagen in C28/I2 cells and up-regulated functional protein expression, which is also functionally important in any consideration of IVD matrix replenishment. Specifically, the mRNA expression of collagen type II, type X and type XXVII was observed, the ramifications of which are not yet clear. Collagen XXVII is a recently discovered fibrillar collagen that is hypothesized to play a role in the organization of cartilage extracellular matrix prior to endochondral ossification.34, 35 Collagen X is also expressed at endochondral ossification, by hypertrophic chondrocytes.36 Interestingly, we have found strong CDMP-2 expression in hypertrophic chondrocytes in foetal spinal tissue,37 and its expression has been linked with the delineation of bone boundaries in developing organisms.28, 29 Further studies are needed to determine the specific signaling pathway of CDMP-2 in regulating expression of specific extracellular matrix molecules both during development and in adult tissue maintenance, but the tendency for CDMP-2 to promote chondrogenic tissue renewal is encouraging from the perspective of therapeutic use in disc regeneration.
Interestingly, when investigating expression of known BMP receptors, we found that BMPR-2 and BMPR-1A were up-regulated by CDMP-2 in C28/I2 cells, suggesting their involvement in CDMP-2 signaling in chondrocytes. Previous functional studies have shown expression of BMPR-2 and BMPR-1A in a CDMP-2 stimulated mesenchymal progenitor cell line,30 but reported a functional preference for CDMP-2 binding to the BMPR-IB/BMPR-2 receptor complex rather than BMPR-IA/BMPR-2.38 There appeared to be no statistically significant up-regulation of BMPR-1B in response to CDMP-2, although increased mRNA expression was measured at 200ng/mL. Interestingly, solid phase protein affinity studies have indicated CDMP-2 has a stronger interaction with BMPR-IB than BMPR-IA.39 With a large number of BMP molecules all utilizing the same receptor complex components to trigger intracellular signaling, receptor usage by BMPs has been proposed as a significant means of controlling the effects of these powerful morphogens.40 Further detailed work will be necessary to understand these factors in disc cells.
While multiple molecules have demonstrated proliferative and chondrogenic effects on existing cells,14, 41 what remains as an obstruction to their use as therapies is the lack of cells in the NP of a severely degenerated disc, making the discovery of a signalling molecule with chemotactic potential for cells residing in the adjacent EP to be an attractive therapeutic option. In biological terms, two aspects must be established in order to illustrate the chemotactic potential of CDMP-2 in the IVD: 1) the ability of CDMP-2 to bind to surface receptors to induce cell motility, and 2) the ability of the target cells, in this case native EP cells (either mature or progenitor), to migrate under a CDMP-2-induced chemotactic gradient. Ex vivo animal IVD models have highlighted the migration of EP cells and bone marrow-derived stem cells under growth factor stimulation.42, 43 This study provides strong supporting evidence for CDMP-2 induced cell motility. Elevated cellular chemotactic markers in response to CDMP-2 seen in our study strongly suggest CDMP-2 induces a metabolically active process of cell migration, possibly mediated by BMPR-2 and BMPR-1A receptors. Moreover, the proliferative and chondrogenic pattern of CDMP-2 on our cell line suggests similar chemotactic and regenerative effects may be observed in native disc tissue.
Notably, the major limitation of this study is the use of an immortalised cell line, with results that cannot be directly extrapolated to native EP chondrocytes. However, cell lines derived from human chondrocytes have been widely used to investigate the mechanisms involved in chondrocytic tissues (e.g. articular cartilage, EP), and have served as models to understand molecular signalling pathways and efficacy of novel therapies.44, 45 In order to investigate the potential ability of CDMP-2 to stimulate disc chondrocyte (EP cells) migration, the cell line we used, C28/ I2, reflects the major characteristics of this particular type of disc cells we wanted to investigate.18 The results from our study provide a rational basis for more focused investigations into the chemotactic actions of CDMP-2 on native discs. Specifically, studies into how CDMP-2 affects the receptor and intracellular pathways of cartilaginous EP cells are worthwhile to confirm the chemotactic potential of CDMP-2 demonstrated in this study.
Conclusion
The current study highlights the ability of CDMP-2 to both stimulate chondrocytes to mobilise and attract cell movement along a chemotactic gradient. In addition, it has reinforced many previous investigations in indicating the chondrogenic potential CDMP-2 in the stimulation of cell proliferation, and synthesis of collagen and proteoglycan at a functional and transcriptional level. Further examination of CDMP-2's biological properties on EP of the IVD both in vivo and in vitro are necessary, which will show a further dimension of possibility for this molecule in application to biological tissue regeneration therapies.
Disclosures
There are no competing interests of any relevant parties involved in this study.
Acknowledgement
The authors would like to thank Spine Service, St. George Hospital, Sydney, Australia for support and funding of this study. In addition we gratefully acknowledge Dr. Mary B. Goldring for the kind gift of the C28/I2 human chondrocytic cell line used in this study. The study was performed at the St George Orthopaedic Research Institute, Sydney.
- Copyright © 2015 ISASS - This manuscript is generously published free of charge by ISASS, the International Society for the Advancement of Spine Surgery
References
In this issue



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Jump to section
Related Articles
Cited By...
- No citing articles found.